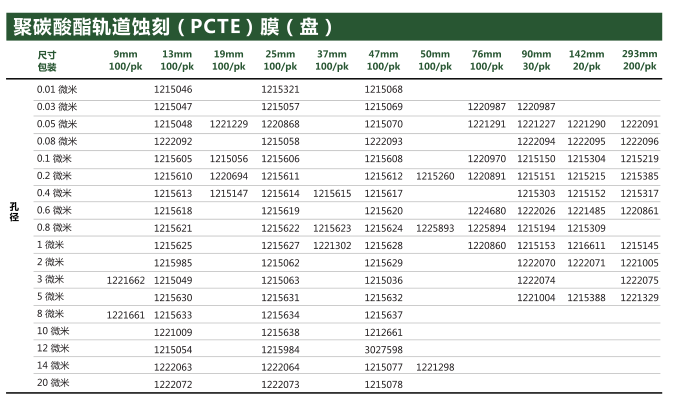
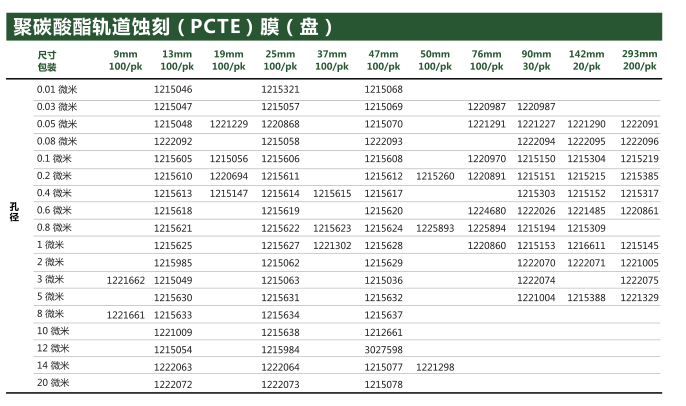
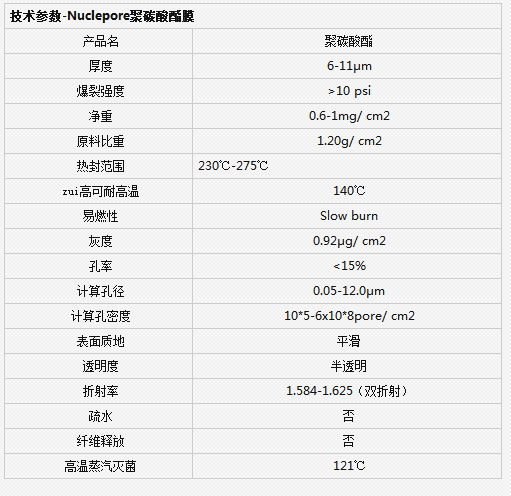
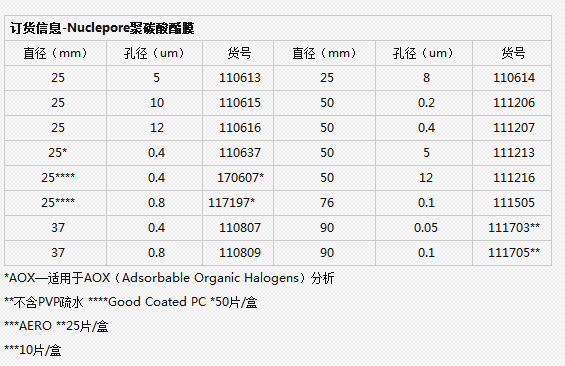
【简单介绍】
【简单介绍】
【详细说明】
原装进口英国Whatman1005-047 Grade 5定性滤纸 GR 5 4.7CM 100/PK
whatman官网 Whatman滤纸 Whatman滤膜

简单介绍:
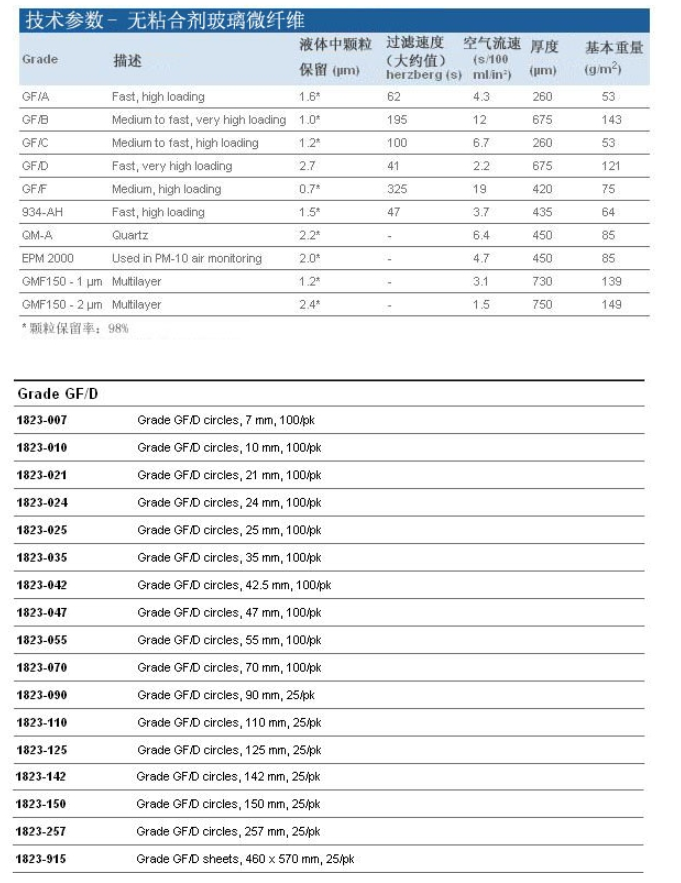
Whatman定性滤纸用于定性分析技术中鉴定物质的性质。折叠好的定性滤纸与相同型号平整的滤纸相比,加快了流速和增加了负载力,定性滤纸―标准级,Wet Strengthened Grades湿强定性滤纸。湿强定性滤纸含有少量的化学稳定树脂,因而增强了湿强度。这不会因此引入任何明显的杂质到滤液中。但因树脂含氮,这些级别的滤纸不能用于凯氏定氮测定。
Whatman定性滤纸的详细介绍:
Whatman定性滤纸Grade1/Grade2/Grade3/Grade4/Grade5/Grade6
Whatman定性滤纸用于定性分析技术中鉴定物质的性质。折叠好的定性滤纸与相同型号平整的滤纸相比,加快了流速和增加了负载力。
定性滤纸―标准级
Grade 1:11μm 在日常过滤中最经常使用的,中等保留力和流速。非常广泛地用于实验室应用,澄清液体。典型地用于沉淀物的定量分析分离,如硫酸铅、草酸钙和碳酸钙。
在农业方面,它用于土壤分析和种子测试;在食品方面,Grade 1滤纸常用于相关液体和抽离液体分离固体食品的常规方法。并且广泛用于教学中简单的定性分析分离。
在空气污染监测方面,有圆片和卷片可供使用,从气流中收集大气灰尘然后用光度计测量;在气体探测中滤纸用发色剂浸湿,用光反射来定量。
Grade 2:8μm 比Grade1保留力略强,但过滤时间却相对长些(即过滤速度相对慢些),吸附性比Grade 1强,已折叠好的为Grade 2V。除8μm大小颗粒的普通过滤处,它额外的吸附性能也派上了用场,例如,在植物生长实验中截留土壤营养,也用于监测空气和土壤测试中的特殊污染物。
Grade 3:6μm 厚度是Grade 1的两倍。负载力好,颗粒保留较小。由于湿强度好适合放在布氏漏斗中使用。其高吸附性能可用作为样品的载体。
Grade 4:20-25μm 非常适用于分析中常规生物液体和有机浸出物澄清的快速过滤,空气污染监测中只要求高流速但对细小颗粒收集要求不严的采样。
Grade 5:2.5μm 最高效的定性滤纸,用于收集小颗粒,流速漫。适用化学分析、澄清悬浮物和水泥土分析。
Grade 6:3μm 流速是Grade 5的两倍,但颗粒保留度相等。常被指定用于锅炉水分析
whatman官网 Whatman滤纸 Whatman滤膜 Whatman中国官网
|
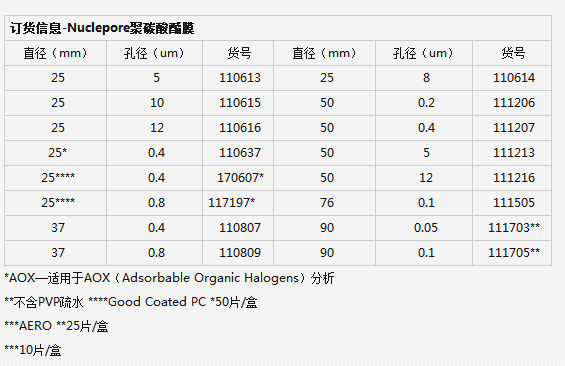
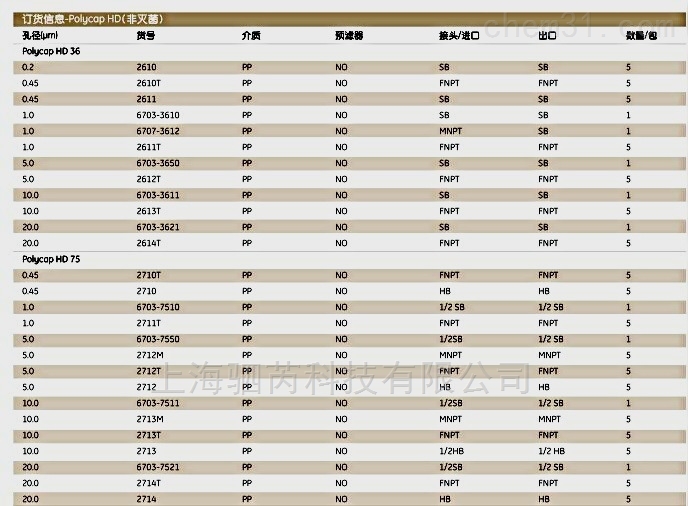
订货信息–定性标准滤纸 |
||||
|
直径(mm) |
Grade 4 |
Grade 5 |
Grade 6 |
数量/ 包装 |
|
10 |
– |
– |
– |
500 |
|
23 |
– |
– |
– |
100 |
|
25 |
1004-325 |
1005-325 |
– |
100 |
|
30 |
– |
– |
– |
100 |
|
32 |
– |
– |
– |
100 |
|
42.5 |
1004-042 |
1005-042 |
1006-042 |
100 |
|
47 |
1004-047 |
1005-047 |
– |
100 |
|
55 |
1004-055 |
1005-055 |
– |
100 |
|
70 |
1004-070 |
1005-070 |
1006-070 |
100 |
|
85 |
– |
– |
– |
100 |
|
90 |
1004-090 |
1005-090 |
1006-090 |
100 |
|
110 |
1004-110 |
1005-110 |
1006-110 |
100 |
|
125 |
1004-125 |
1005-125 |
1006-125 |
100 |
|
150 |
1004-150 |
1005-150 |
1006-150 |
100 |
|
185 |
1004-185 |
1005-185 |
1006-185 |
100 |
|
240 |
1004-240 |
1005-240 |
1006-240 |
100 |
|
270 |
1004-270 |
– |
– |
100 |
|
320 |
1004-320 |
1005-320 |
– |
100 |
|
400 |
1004-400 |
– |
– |
100 |
|
500 |
– |
– |
– |
100 |
|
FilterCup 70* |
– |
– |
– |
25 |
英国沃特曼一级代理_whatman总代理_玻璃微纤维滤纸_纤维素层析纸
whatman官网 Whatman滤纸 Whatman滤膜
*一次性购买带橡胶塞过滤杯底座–货号1600-900
上海金畔生物科技有限公司
文章号20207688-20207688












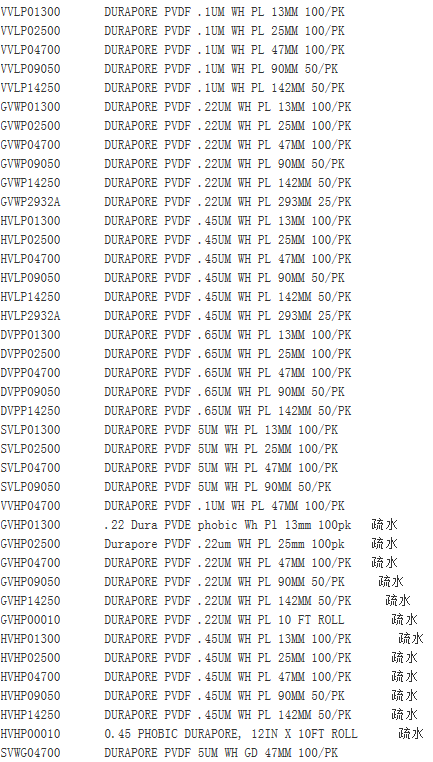

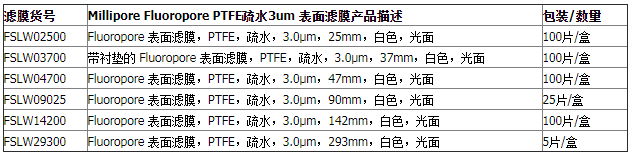
 Millipore孔径0.45um聚四氟乙烯疏水滤膜 FHLP04700
Millipore孔径0.45um聚四氟乙烯疏水滤膜 FHLP04700